Top 10
Prehistoric
Marine Predators
#10 – DUNKLEOSTEUS
Phonetic: Dun-kel-os-tee-us
Named By: Lehman - 1956
Synonyms: Dinichthys terrelli, Ponerichthys
Classification: Chordata, Placodermi, Arthrodira, Dunkleosteidae
Species: D. terrelli (type), D. amblydoratus, D. marsaisi, D. raveri
Diet: Carnivore/Piscivore
Size: Up to 10 meters long
Known locations: Canada. USA. Europe. Morocco
Time period: Famennian of the Devonian
Fossil representation: Several specimens, but only the bony head and jaws are known
In modern popular culture, Dunkleosteus is by far the best known and most often represented of the early placoderm carnivores. However, Dunkleosteus actually sat within the Dinichthys genus for a long time as the species Dinichthys terrelli. It was not until the large numbers of Dinichthys remains were re-studied that it was realized that a large number of the Dinichthys fossils actually represented different genera, not species. The result was that many of these remains were split to form new genera including the creation of Dunkleosteus. In a further twist however, the species that was split to form Dunkleosteus, D. terrelli, was actually the remains most often used when reconstructing Dinichthys. Along with the equally giant Titanichthys, Dunkleosteus is one of the largest predatory placoderm fish known in the fossil record. However unlike Titanichthys which seems to have preferred large quantities of smaller prey items,Dunkleosteus was an apex predator capable of taking down pretty much anything it could clasp its jaws around.
Because only the armored head of Dunkleosteus is known, it can be problematic to reconstruct the rear portion that was almost certainly unarmored. This reasoning comes about from the lack of rear fossils for not just Dunkleosteus but all related placoderms that are similar to Dunkleosteus. The only possible insights come from much smaller placoderms like Coccosteus. With a maximum length approaching only forty centimeters, Coccosteus was positively tiny in comparison to Dunkleosteus, but the impressions of the softer hind body can still be seen in fossils attributed to it. Smaller animals in general tend to fossilize better and in more complete states because they can get protected from scavengers and the elements more quickly. Aside from armor, the bony plates of Dunkleosteus could have served two further purposes.
























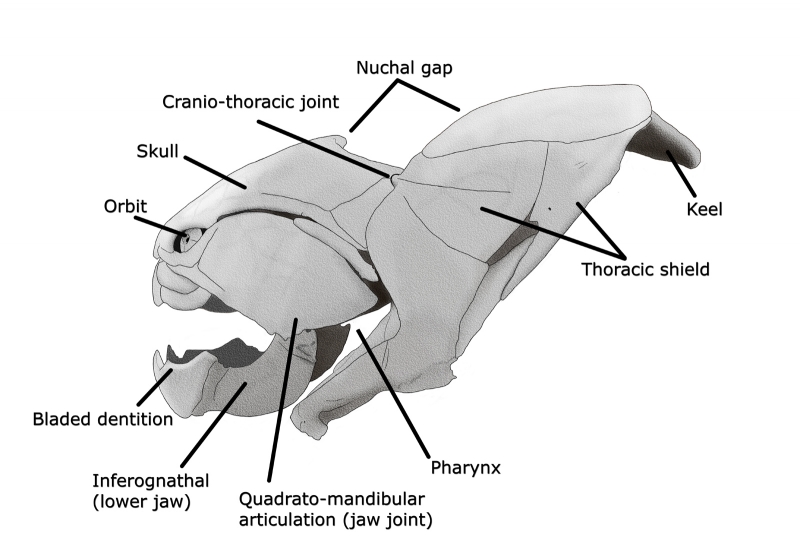
As a predatory placoderm, Dunkleosteus likely attacked other related placoderms that had the same kind of bony plates for protection. In the absence of a harder organic material, Dunkleosteus would at least need the same material to break through the armor. Shape the material to a sharp bladed edge, and you have a chance of cutting through it. The second reason is that the jaws need enough driving power to cut and break apart armored prey, and the only way that this could happen is if Dunkleosteus had incredibly powerful jaw muscles. However these muscles would in turn require strong supports and attachments otherwise the jaws could not be brought to bear with their full force. This is likely why Dunkleosteus and other related placoderms like Eastmanosteus retained armored heads. Although Dunkleosteus had a powerful bite it was not the strongest among fish, that title goes to the gigantic shark Megalodon that lived several hundred million years later. Still, with a bite force estimated at almost two metric tons,Dunkleosteus could still use its sharp jaws to shear through any prey item it chose. What prey items were on the menu seem to have been dependent upon the age of the Dunkleosteus in question. Juvenile Dunkleosteus had stiff jaws best suited for soft-bodied prey items. As Dunkleosteus grew older, the jaws would become more flexible, perhaps to better protect them from injury and breakage when dealing with larger, more powerful and more heavily armored prey items. The actual hunting and feeding style of Dunkleosteus is also a popular subject of interest. With the heavy plates around the head, Dunkleosteus was probably not a fast swimmer, but still would have had powerful muscles developed from just swimming around with the weight. This meant that Dunkleosteus either preferred slower prey, or used ambush tactics to try to take its prey off guard. Another thing to consider is that the jaws of Dunkleosteus, and probably other similar placoderms, could open exceptionally fast within a fraction of a second. This would create a sudden void inside the mouth of Dunkleosteus, creating a vacuum that sucked the water and the prey that was swimming in it into its mouth. This means that Dunkleosteus did not have to physically catch its prey, just get close enough to open its mouth.
Larger Dunkleosteus seemed to have preferred other placoderm fish perhaps like Bothriolepis that was very common at the time. Once caught, Dunkleosteus could use its sharp jaws to cut up prey, but it seems that only the softer and more easily digestible flesh was desired. When Dunkleosteus fossils are found, they are often found in association with fish boluses. A bolus is a ball of remains that has been chewed and swallowed, but the remains in connection with Dunkleosteus are of only bones that already seem to have been partially digested. This indicates that Dunkleosteus may have spat out parts that were too hard for it to digest completely, a precedent that is known in numerous other fish. Finally, because Dunkleosteus was at the top of its food chain, the only other thing that it would have to worry about being attacked by was another Dunkleosteus. This actually does seem to have happened with some Dunkleosteus plates actually showing damage that seems to have been inflicted by the jaws of another Dunkleosteus. Asides from territorial combat, this may indicate active cannibalism in Dunkleosteus.
Phonetic: Shas-tah-sore-us
Named By: Merriam - 1895
Synonyms: Shastasaurus alexandrae, Shastasaurus osmonti, Guanlingsaurus liangae, Shonisaurus sikanniensis
Classification: Chordata, Reptilia, Ichthyosauria, Shastasauridae
Species: S. pacificus (type), S. liangae, S. sikanniensis
Diet: Specialist of soft-bodied cephalopods like fish
Size: 21 meters long for S. sikanniensis, other species like S. liangae seem to be 4 to 9 meters long based upon available fossils
Known locations: USA, California - Mount Shasta. Canada, British Columbia - Pardonet Formation. China
Time period: Carnian to Norian of the Triassic
Fossil representation: Many specimens, although often incomplete and fragmentary
Shastasaurus hit the record books in 2011 with the re-examination of a set of ichthyosaur remains from British Columbia that were initially described as a new species of Shonisaurus back in 2004. Study of the remains has found that while they are similar, the remains are a better match for Shastasaurus and so the remains were moved from Shonisaurus to create a new third species,Shastasaurus sikanniensis. This confusion is not surprising however when you consider that these two ichthyosaurs are thought to be very similar to each other in not just form but predatory behavior as well. Although the material referable to S. sikanniensis has allowed for a new upper size of the genus to be established, it is Chinese fossil material referable to the second species, S.liangae named in 2000 that has allowed for considerably more accurate reconstructions. The type species of Shastasaurus was only ever known from fragmentary remains with additional fossils being attributed to it later. More complete S. liangae material however revealed that Shastasaurus is a short-snouted ichthyosaur and as such much of the long-snouted ichthyosaur fossils associated with the type species were found to have actually been placed in the wrong genus, and have since been moved to other genera.
Today, Shastasaurus remains are usually associated with the Northern coast lines of the Pacific Ocean, but it needs to be remembered that back in the Triassic today’s continents were still very close to one another and surrounded by a single large ocean. While the Pacific Rim would have formed much of this coastline, the gradually breaking up land masses created new seas and it is thought by some paleontologists that Shastasaurus may have ventured into these new areas as well. Shastasaurus also has two dubious species associated with the genus, S. carinthiacus from Austria and S. neubigi from Germany. Central Europe at the time of the Triassic was more like an Island chain surrounded by shallow seas, not only well away from other Shastasaurus fossil sites, but different from the deep water environments where Shastasaurus is thought to have hunted. If these remains are not those of Shastasaurus then they may yet represent a similar type of ichthyosaur, whereas if they are the same genus then they may reveal that Shastasaurus was either more flexible in its hunting, or that it was possibly migratory and heading to shallow water birthing areas or moving on to new feeding grounds.
















Shastasaurus was what is now termed a primitive ichthyosaur. Key to this is what appears to be a lack of a dorsal fin, something which is clearly seen in advanced ichthyosaurs but is lacking in basal forms. The lack of a dorsal fin bone does not necessarily mean that a dorsal fin was not present; it may have existed in the form of softer tissue such as cartilage which rapidly decomposes in sea water. Still without more direct evidence the wider assumption is that Shastasaurus did not have a dorsal fin. The tail fluke of Shastasaurus may not have been as well-formed as in later ichthyosaurs either. The rear of the body that grew into the tail is seen to have strongly arced downwards indicating that the lower tail lobe was larger and more developed than the upper lobe. This suggests that Shastasaurus was better at cruising rather than active pursuit of prey.
Early art depicting ichthyosaurs often depicts them to have large rounded bodies, and while modern reconstructions suggest that ichthyosaurs in general are much slimmer than what they were classically depicted to be, the ribs in proportion to total body length prove that this was definitely not the case for Shastasaurus. The distance between fore and aft flippers is roughly 7 meters while the ribs were barely 2 meters tall. This gives Shastasaurus a profile more like that of a torpedo which would have helped to streamline Shastasaurus as it swam through the water. This effectively means that Shastasaurus faced reduced resistance and experienced less drag so that it did not have to expend so much energy on swimming, something that is especially important given the underdevelopment of the tail fluke. As mentioned above, the fossil specimens of S. liangae from China allowed for the first accurate reconstruction and analysis of Shastasaurus, particularly the skull. The skull itself is just a little bit over eight per cent of the total length of adult Shastasaurus, and displays extreme shortening adaptations. While the jaws are reduced, they seem to have reached their minimization limit as the nasal bone which is usually mid-way in the skull is on the very tip of the jaws. The skull is also laterally compressed which means that when viewed from the front the skull would look narrow like it was pushed in from the sides.
The addition of new fossil material to create the species S. sikanniensis has made Shastasaurus the largest currently known marine reptile at twenty-one meters long. This is much larger than others such as the Jurassic predator Liopleurodon which is often incorrectly labeled as the largest and credited as being up to 25 meters long, even though the largest fossil specimens for this genus are barely 6.5 meters long. Shastasaurus is thought to have grown to such a large size so that it was better able to dive to deep depths, as well as being larger than most of the marine predators of the time effectively taking full-grown adults off the menu list. Large size meant that it could take in a greater amount of air and stay down longer than a much smaller ichthyosaur. Shastasaurus has one of the most specialized mouths of all known ichthyosaurs in that the jaws are both short and toothless. This toothless adaptation is also seen in the similar Shonisaurus although this ichthyosaur has comparatively longer jaws. Why Shastasaurus should have such short jaws has puzzled many, but analysis of the skulls, particularly the attachments for the jaw opening muscles has given rise to the theory (proposed by M.P. Sander, X. Chen, L. Cheng & X. Wang in 2011) that Shastasaurus was a suction feeder. Like some fish Shastasaurus opening its mouth wide so fast that it would have created a point where the space and existing water inside its mouth would be reduced to such a low pressure it would create a vacuum inside its mouth. Water immediately in front of the mouth, as well as anything in it such as squid, would get sucked into this vacuum as the pressure within was equalized.
Further evidence for this feeding behavior comes from the large hyoid bones that would have allowed the tongues to have also been quickly drawn low and back, further increasing the vacuum effect. The very size of the jaws themselves also support this as shorter jaws would have much less resistance when moving through the water than longer jaws, further increasing the speed that they can be opened which in turn increases the vacuum effect inside the mouth. If this proposed method of feeding is correct then it’s a wonderful case of nature reverse engineering itself. This is because squid use jets of water pressure squirted out of their bodies to propel themselves through the water, especially when escaping predators. Shastasaurus on the other hand basing its feeding behavior on using the hydro dynamics of an aquatic vacuum would have denied the squid their advantage, making prey capture for Shastasaurus quite easy.
Phonetic: El-lazz-mo-sore-us
Named By: Edward Drinker Cope - 1868
Classification: Chordata, Reptilia, Sauropterygia, Plesiosauria, Elasmosauridae
Species: E. platyurus (type)
Diet: Piscivore
Size: 14 meters long
Known locations: USA, particularly associated with Kansas - Pierre Shale
Time period: Cretaceous
Fossil representation: Many specimens
Today Elasmosaurus is one of the best known plesiosaurs in the world and is the type genus for the Elasmosauridae. This group is noted for having particularly long necks, proportionately even longer than the more standard plesiosaurs. These long necks are composed of a large number of cervical vertebrae that in Elasmosaurus number 71, more than other plesiosaurs including fellow elasmosaurids. The sheer number of vertebrae not only extend the length of the neck but also provide greater flexibility for fine control of the head, although very early reconstructions of Elasmosaurus being able to coil its neck like a snake are today considered to be very inaccurate and impossible for a living Elasmosaurus to do. Instead the neck would have been held quite straight when resting with side-ward movements only occurring when necessary.
It was the long neck of Elasmosaurus that was key to its feeding method. All Elasmosaurus would have to do was swim up to a shoal of fish, possibly from below so that it could hide its body in the slightly darker depths, and use its neck to dart its head in and pluck out a mouthful of fish. Approaching prey like a shoal of fish from below would also silhouette themselves against the brighter surface water, making them easier to define as well as removing the defense method of flashing light off their shiny scales to confuse predators as they turned and swam through the water. Such a method would have been a reasonably efficient form of feeding as Elasmosaurus would just have to move its head and neck and not waste energy swimming after fast-moving individuals. Key to prey capture were the long and thin teeth that protruded from the mouth of Elasmosaurus. These inter-meshed together so that when a fish was caught between the jaws it was impaled upon the teeth so that it could not wriggle free & escape. This kind of dentition is common among other elasmosaurids as well as other piscivorous (fish-eating) animals such as skim feeding pterosaurs. A defining characteristic of Elasmosaurus is the fact that it has six teeth in each premaxilla. Because the teeth of Elasmosaurus were adapted for prey capture rather than biting clean through flesh, prey would need to have been swallowed whole. Gastroliths have been found in association to Elasmosaurus remains and its thought that the grinding action of these stones in its gut tenderized food for easier digestion.



























Elasmosaurus probably was not a fast swimmer, although if it used stalking tactics to approach its prey from below it would not need to be. The flipper-like limbs were rigid and well suited for providing efficient locomotion through the water. Skeletal elements of the main body such as the pectoral girdles were also well adapted to provide efficient musculature support for the flippers so that Elasmosaurus could paddle itself through the water with ease. While Elasmosaurus was beautifully adapted for an aquatic life, the price of these adaptations is that Elasmosaurus would have almost certainly been restricted to the marine environment. Because the paddle-shaped limbs were so stiff they would at best been only just better than useless for pushing its body along. Even when if Elasmosaurus was ever active in the shallows, it would still need to have kept its body totally immersed so that the water could support its body weight.
More classic depictions of Elasmosaurus shooting its neck out of the water are also unlikely as the longer the neck got, the more body tissue such as bone and muscle was used to make up the neck meaning more weight was added to it. More weight means that stronger muscles would have been needed to support the neck when out of the water and study of the cervical vertebrae where the muscles attach indicates that Elasmosaurus just did not have the muscles to move its neck when it did not have water to support the weight. Instead the muscles were only strong enough to provide basic neck movements, which would be all that was required of them when the neck was immersed in water. While no hard evidence currently exists for Elasmosaurus either laying eggs on land or giving birth to live young, most paleontologists believe that Elasmosaurus and similar marine reptiles would have given birth to live young. The precedent is known to exist for other marine reptiles like the Ichthyosaurs as far back as the Triassic and given the problems that Elasmosaurus would have had just moving its body if it ever found itself out of the water make it extremely unlikely it could have laid eggs on land. Today marine reptiles such as sea snakes can also be observed giving birth to live young which indicates that as a group the reptiles are capable of adapting to new methods to thrive in different environments.
The palaeontological history of Elasmosaurus is a turbulent one. First discovered in 1868 by Dr. Theophilus Turner, the fossils were sent to Edward Drinker Cope for study and identification. Cope had never assembled a plesiosaur before but he had the idea the large number of vertebrae mostly belonged in the tail which resulted in a long tail and short neck. To Cope this made sense as the proportions were similar to those of many modern lizards that also have short necks and incredibly long tails. The next part of this story is a little hazy for exact details, but the popular retelling of it has Othniel Charles Marsh realizing that the head was on the wrong end and ridiculed Cope for placing it in the wrong place. However it seems that it was actually Joseph Leidy who pointed this error out in a meeting in 1870, although given the animosity already between Marsh and Cope at the time, its quite possible that Marsh continued to use this case as an example to discredit Cope’s work, hence the confusion. It is this criticism that poured fuel onto the already burning fire that would become the “bone wars”, a fierce rivalry between Cope and Marsh that saw them trying to out compete one another in terms of discovering new species. Since its naming back in 1868, many new specimens of Elasmosaurus have been recovered and many of these were assigned their own species within the genus. Many of these species continue to be mentioned despite the fact that in 1999 the paleontologist Ken Carpenter conducted a study of the known Elasmosaurus fossils at this point and found that some of these did not belong as individual species and some of them even represented different genera. Today most paleontologists only recognize the original type species, E. platyurus as being valid.
Phonetic: Krow-no-sore-us
Named By: Longman - 1924
Classification: Chordata, Reptilia, Sauropterygia, Plesiosauria, Pliosauroidea, Pliosauridae
Species: K. queenslandicus (type), K. boyacensis
Diet: Carnivore
Size: Estimated between 9 and 10 meters long. Skull averages 2.7 meters long
Known locations: Australia
Time period: Aptian to Albian of the Cretaceous
Fossil representation: Two sets of remains and associated individual teeth
Kronosaurus is one of the more famous fossil discoveries to come from Australia and as a huge short-necked pliosaur, Kronosaurus would have been an apex predator of the waters it swam in. The teeth of Kronosaurus are conical and up to thirty centimeters long, although most of that was root with up to twelve centimeters making up the exposed crown. Interestingly the teeth of Kronosaurus are quite different to other pliosaurs in that they are not serrated and do not have three facets. The teeth may suggest that despite its large size Kronosaurus may have seized smaller to medium-sized prey which could then be manipulated to be swallowed. The only way that Kronosaurus could have fed from larger prey would be to clamp on with its jaws and then use brute strength from its body to tear off a chunk of flesh by twisting or rolling. Either way it seems that Kronosaurus may have eaten in a similar manner to a crocodile.































One prey type for Kronosaurus seems to have been the long-necked elasmosaurid plesiosaurs that were quite common in the southern oceans of the Cretaceous. Evidence for this predation comes from the skull of an Eromangasaurus that seems to have Kronosaurus bite marks. Also the fact that Eromangasaurus is only known from a skull might suggest that the head was torn off in the Kronosaurus attack. Initial size estimates for Kronosaurus placed it at just under 13 meters long. However further study combined with a better understanding of pliosaur remains in general resulted in a reduction in this estimate to somewhere in the region of nine to ten meters long. This estimate is universally accepted as being more accurate, and although the upper size is reduced to ten meters, Kronosaurus was still big for a pliosaur and was much bigger than Liopleurodon which is often incorrectly credited as the biggest pliosaur ever. An even bigger pliosaur than Kronosaurus however is the one that has been dubbed ”Predator X”.
#6 – BASILOSAURUS
Phonetic: Ba-sil-oh-sore-us
Named By: Richard Harlan - 1847
Synonyms: Zeuglodon
Classification: Chordata, Mammalia, Cetacea, Archaeoceti, Basilosauridae
Species: B. cetoides (type), B. drazindai, B. harwoodi, B. isis, B. caucasicus, B. paulsoni, B. puschi, B. vredensis, B. wanklyni
Type: Carnivore
Size: Average 18 meters long
Known locations: USA. Egypt, Wadi Al-Hitan. Pakistan
Time period: Bartonian to Priabonian of the Eocene
Fossil representation: Lots of known specimens
The one thing about Basilosaurus that instantly causes confusion is its name. When first studied and named by Richard Harlan, he came to the conclusion that it was most likely a marine reptile and so named it Basilosaurus or “King Lizard”. It was not until the remains were studied by Sir Richard Owen that they were confirmed as mammalian. Owen then proposed that Basilosaurus should be renamed Zeuglodon (Yoke Teeth) but because Basilosaurus was the officially registered name, it could not be changed, so Zeuglodon became a synonym instead of a replacement. Basilosaurus represents one of the earliest whales although it’s actually descended from terrestrial mammals like Ambulocetus. Evidence for this can be seen in the flippers. The front flippers still have an elbow joint, something that today is only seen in seals. The back flippers where the hind legs would have been in its ancestor are greatly reduced and although they may have been used to get extra grip on a mate, they would eventually disappear in later whales. The size of its flippers in comparison with the massive bulk of its body means that it was almost certainly an entirely pelagic animal. The large jaws of Basilosaurus housed teeth suitable for catching prey that would not have been especially small, perhaps smaller whales or large fish that while smaller than Basilosaurus, but were still too large for most other predators to tackle. Superficially, the fossils of Basilosaurus skulls resemble the skulls of mosasaurs that swam the oceans tens of millions of years earlier, disappearing from the fossil record at the time of the KT extinction 65 million years ago. It is partly for this reason that Basilosaurus was misidentified as a reptile because mosasaur remains were known, yet no one was aware that the early whales looked so similar.











































The skull also has a reduced area for the brain when compared to modern whales. Going on the basis that larger brained cetaceans are typically more social, Basilosaurus itself may have been a solitary predator. Although not preserved, the structure of the tail vertebrae suggests support for a tail fluke as seen in modern cetaceans. The reduced limbs would probably have been of little use in actual locomotion leading to the suggestion that Basilosaurus used an undulating motion to propel itself in the water. This up and down motion may have also provided quick bursts of speed at prey items. Study of the Basilosaurus skeleton has revealed that it was quite restricted in terms of movement. Muscle attachments along the spine imply that Basilosaurus had relatively weak muscles and could neither dive deep nor swim for extended periods, at least at what may have been pursuit speed for prey. The vertebrae, unlike modern whales which are solid, were hollow and likely filled with fluid. This is not an adaptation for a deep-sea creature, as pressure imbalance between the fluid in the hollow vertebrae and stronger pressure of deep water outside could in theory cause spinal injury with the vertebrae being crushed if the pressure became too much. Together, these all point to an animal that only swam and hunted in waters near the surface. In 1845, Dr. Albert Koch constructed a composite animal from numerous specimens of Basilosaurus remains. This composite was named Hydrarchos and went on a touring display around the world until its destruction in the Great Chicago fire in 1871. Aside from never existing, the material was presented as a sea serpent not a whale. Also not only was their more than one Basilosaurus individual used in the construction, fossils from other animals were also included. Despite the fact that other scientists of the time were quick to point out that it did not represent a real creature, references to it as a real animal are still sometimes made by those who are not knowledgeable about this history, although thankfully this is getting rarer.
#5 – CRETOXYRHINA
Phonetic: Creh-tox-ee-rye-nah
Named By: Louis Agassiz – 1843
Classification: Chordata, Chondrichthyes, Elasmobranchii, Selachimorpha, Lamniformes, Cretoxyrhinidae
Species: C. mantelli
Type: Carnivore
Size: Up to 7 meters long
Known locations: Worldwide
Time period: Cenomanian to Campanian of the Cretaceous
Fossil representation: Usually teeth, but some impressions revealing the cartilaginous skeleton are also known
Also known as the “Ginsu Shark”, Cretoxyrhina was the biggest shark of its time, considerably bigger even than its contemporary Squalicorax. Cretoxyrhina was near the top of the food chain and it had a diet that included almost anything that was in front of it, including mosasaurs. There is also strong evidence for its predation of the bony fish Xiphactinus, a 4 to 6 meter long predator itself and even the large turtles of the day were fair game for Cretoxyrhina. This species acquired the name “Ginsu Shark” in reference to its teeth. At up to seven centimeters long, the teeth are smooth and have thick enamel, suggesting that they were for work against tough shells and bones. Like the advert for Ginsu knifes said, they could cut through anything and Cretoxyrhina probably had little difficulty in removing bite sized chunks from whatever prey item it chose. The overall morphology for Cretoxyrhina is lamniform, similar to a Great White Shark today. The tail also shows adaption form fast swimming, enabling Cretoxyrhina to give chase after faster prey.























During the late Cretaceous about 100 million years ago, there arose a ferocious expression of lamnoid predatory prowess named Cretoxyrhina. Growing to a length of 15 to 18 feet (4.5 to 5.5 meters), Cretoxyrhina rivaled the modern Great White Shark in sheer size. Most Cretaceous sharks are known primarily from isolated fossilized teeth. In contrast, several nearly complete skeletons of Cretoxyrhina have been discovered. These astonishing fossil finds include dis-articulated clusters of vertebrae with associated upper and lower tooth sets lying in clear arcs, giving a vivid impression of Cretoxyrhina‘s body length and mouth width. Remarkably, paleontologists have also discovered at least 2 mineralized impressions of Cretoxyrhina skin, with tooth-like dermal denticles still anchored in place. From these and other, less spectacular finds, we know that Cretoxyrhina lived globally in oceanic and coastal areas, including the Western Interior Sea (sometimes known by more evocative name, Sundance Sea) which had flooded inland North America during the Mesozoic.
#4 – PLIOSAURUS – a.k.a. “Predator X”
Phonetic: Ply-oh-sore-us
Named By: Richard Owen - 1841
Synonyms: Liopleurodon rossicus, Plesiosaurus giganteus, Plesiosaurus recentior, Spondylosaurus, Stretosaurus, Strongylokroptaphus
Classification: Chordata, Reptilia, Sauropterygia, Plesiosauria, Pliosauridae
Species: P. brachydeirus (type) P. funkei, P. kevani, P. macromerus, P. portentificus. Additional species include P. brachyspondylus, P. carpenteri, P. macromerus, P. portentificus, P. rossicus and P. westburyensis though these are not always recognised as valid
Diet: Carnivore
Size: Some species up to 10 meters long, with skulls approximately 2 meters long Largest species, P. funkei estimated at between 10-12.8 meters long
Known locations: England - Kimmeridge Clay Formation, and Norway, Svalbard, Spitsbergen. Possibly also France, Poland and Russia, depending upon the validity of all species
Time period: Kimmeridgian to Tithonian of the Jurassic
Fossil representation: Numerous specimens
Originally described as a species of Plesiosaurus, this was the first pliosaur ever named which is why Pliosaurus is used as the type genus of the Pliosauridae. Related to the long-necked plesiosaurs the pliosaurs adapted to fill a range of different predatory niches with some like Pliosaurus itself being hunters of other marine reptiles. Such prey preference is indicated by the large teeth as much as thirty centimeters long of the jaws which would have been okay on small prey like fish, but devastating against large prey like plesiosaurs, ichthyosaurs, smaller pliosaurs and quite possibly giant fish such as Leedsichthys. Because of its long history a large number of remains under many different species have been attributed to the genus, but the list of valid names has now been greatly reduced as a result of more intensive study of the fossils. The earlier attribution of fossils under multiple species names is known as the wastebasket effect and is something that happened to almost all early discovered prehistoric creatures including the dinosaurs Megalosaurus and Iguanodon as well as the pterosaurs Pterodactylus and Rhamphorhychus.



































In 2006, highly fragmentary remains of a marine reptile were found and in 2008, an effort began to recover the fossils from the island of Spitsbergen in the Svalbard archipelago. These fragments numbered around 20,000 pieces in number and took a long while to piece together though like a 3D jigsaw, they slowly came together to form a particularly huge Pliosaur. “Predator X” quickly caught the attention of the news and popular science media, and was even a focus animal in an episode of the well-made dinosaur documentary series Planet Dinosaur, all before this individual had even been granted an official binomial name. Today, ”Predator X” is known within scientific circles as Pliosaurus funkei which is the official descriptive name for this pliosaur that was granted in 2012. When P. funkei was still only known as Predator X, it was initially thought to have possibly been as much as 15 meters long. Now that the fossils of this pliosaur have been reconstructed more, studied and identified as a species of Pliosaurus, more accurate reconstructions have now been put together. This has seen the speculated size of Predator X, now as Pliosaurus funkei, estimated at somewhere between 10 and 12.8 meters long. Unfortunately this revision has still gone unnoticed by some who still credit individuals as being 15 meters long, in a similar manner to how another pliosaur named Liopleurodon is still being credited as being 25 meters long by some even though the largest known specimen is a little over 6 meters in length. At the time of writing the only known fossils of Pliosaurus funkei include a partial skull, a flipper and a few additional post cranial remains such as vertebrae. These remains are what we’re used to establish the new size ranges for this pliosaur, though it is quite conceivable that should a second specimen ever be found the size range may be revised to be smaller or even a little larger to what it currently stands at. Even at ten meters long, however, P. funkei will still be comparable to other large species of Pliosaurus, as well as other large pliosaur genera such as Kronosaurus and Brachauchenius. Like with its relatives, individual P. funkei would have been specialized predators of other large marine organisms.
#3 – LIVYATAN
Phonetic: Liv-ya-tan
Named By: Lambert et al. - 2010
Homonym: Leviathan melvillei
Classification: Chordata, Mammalia, Cetacea, Physeteroidea
Species: L. melvillei (type)
Diet: Carnivore
Size: Skull is 3 meters long. Total size estimates vary between 13.5 and 17.5 meters long
Known locations: Peru - Pisco Formation
Time period: Serravallian stage of the Miocene of the Neogene
Fossil representation: Partial preserved skull, mandible (lower jaw) and teeth
When first described, Livyatan was actually named Leviathan after the biblical sea monster. Unfortunately however, Leviathan had already been used to name a mastodon now known as Mammut (Leviathan is actually a synonym to this genus, but still cannot be used). As such the Hebrew word for Leviathan, Livyatan, is now used to refer to this ancient whale. The species name, L. melvillei is in honor of the author Herman Melville, the man who wrote the world-famous novel “Moby Dick”. Because only the skull is known, Livyatan is often compared to the Sperm Whale (Physeter macrocephalus) when piecing together the rest of the body. This has yielded size estimates of thirteen and a half meters which make Livyatan a comparable size to the mega-toothed sharks like Megalodon. However Livyatan has also been compared to another prehistoric whale named Zygophyseter and this resulted in an estimate of 17.5 meters. If accurate then this would make Livyatan comparable to even Megalodon, the largest shark to ever swim the ocean. In fact for a time, both Livyatan and Megalodon would have swam in the same oceans. Not only was Livyatan an apex predator like Megalodon, it probably fed upon the same prey animals which were medium-sized baleen whales. Smaller juvenile Livyatan may have preferred proportionately smaller prey items like the smaller cetaceans or even large fish. With teeth that were 36 centimetres long, Livyatan had the dentition to take down large prey items. In fact not only were the teeth of Livyatan considerably larger than Megalodon teeth, they are considered to be the largest known teeth for the purpose of eating.






















Granted some animals like Elephant have modified teeth that form tusks which are even bigger, but they are useless for processing food in the mouth so in this case they do not count as the largest. Livyatan also had teeth in both the upper and lower jaws for grasping prey (the modern Sperm Whale, the closest living analogy to Livyatan, only has teeth on the lower jaw). How Livyatan hunted it still a matter of debate, but given its large mouth and teeth it may have used a similar method to kill smaller whales as Megalodon did. This could be approaching from the bottom and slamming into its target from underneath. An associated method could also be trapping the smaller whale’s rib cage in its jaws and crushing the ribs to create fatal injuries to the internal organs. Another method could see Livyatan holding down a whale beneath the surface to stop it surfacing for air. This is a strategy that would be potentially risky for Livyatan as it too would still need to surface to breathe air, but assuming Livyatan could hold its breath for longer than its prey, it would still be a viable strategy. Livyatan is morphologically similar to the modern Sperm Whale and this has brought comparisons between the two for head function. Livyatan is thought to have had a spermaceti organ that would have been filled with wax and oil. This has also inferred the possibility that Livyatan may have used echolocation to find its prey.
#2 – TYLOSAURUS
Phonetic: Tie-lo-sore-us
Named By: Othniel Charles Marsh - 1872
Classification: Chordata, Reptilia, Squamata, Scleroglossa, Anguimorpha, Mosasauridae, Tylosaurinae
Species: T. prioriger (type), T. capensis, T. kansasensis, K. nepaeolicus, K. pembinensis, K. saskatchewaensis
Diet: Carnivore
Size: Up to 15 meters long
Known locations: Canada. USA, particularly the mid-eastern central states
Time period: Maastrichtian of the Cretaceous
Fossil representation: Many specimens as well as numerous teeth
The discovery of Tylosaurus harks back to the “bone wars”, a fierce rivalry between the palaeontologists Edward Drinker Cope and Othniel Charles Marsh that took place towards the end of the nineteenth century. As such the naming history of Tylosaurus is muddled and can cause confusion to those who are not aware of the full history of Tylosaurus’s naming. The first specimen of a skull and vertebrae discovered in Kansas in 1868 was given the name Macrosaurus prioger by Cope. Approximately a year later Cope re-assigned the material to Liodon. Marsh described a more complete individual in 1872, however his name Rhinosaurus (Nose Lizard) had already been used, and its replacement Rhamposaurus (Beak Lizard) was also already in use by another animal. Both of these proposed names referenced the strong snout and this is loosely reflected in the final name of Tylosaurus (Knob Lizard). This included all of his specimens as well as the material named by cope to produce the type speciesT. prioger. There are a large number of fossils associated with Tylosaurus, mostly from the central United States which was once the sea floor of the Western interior Seaway, but also in other areas of North America. This has resulted in a great number of species being erected for the genus, although as often proves the case with prehistoric animals whose discovery dates back to the nineteenth century, many of these have been found to be synonymous with existing older species as well as other mosasaur genera. This is why modern listings of Tylosaurus usually have much smaller species listings than older ones. One often mentioned species of Tylosaurus, T. haumuriensis described from material in New Zealand has since been declared to be a synonym of theTaniwhasaurus type species T. oweni.Tylosaurus was one of the larger mosasaurs that lived towards the end of the Cretaceous period, something which has secured its frequent inclusion in popular media such as books and television documentaries. Rivals to Tylosaurus in terms of upper size include Mosasaurus and Hainosaurus. Unlike the earlier pliosaurs which relied upon their flippers for locomotion, mosasaurs relied upon their tails to propel themselves though the water and Tylosaurus was certainly no exception. The tail consisted of more that eighty vertebrae, each with a tall neural spine that projected upwards and a deep ’V’ shaped chevron on the underside. These vertebrae resulted in a tail that was laterally compressed but deep so that the maximum surface area available could be used for pushing against the water and propelling Tylosaurus forward. Although not entirely fusiform like a fish, the overall body shape was adapted to be as hydrodynamic as possible. When viewed from the side Tylosaurus was a very big animal, but when viewed from the front Tylosaurus would have presented a very small cross-section. This, combined with the long sloping skull, resulted in a large reduction in the amount of resistance that Tylosaurus experienced as it swam through the water. Also the flippers that were proportionately smaller than those found in pliosaurs were used more for steering than actual swimming, again resulting in a reduction of drag.









































The specialized and occasionally damaged snout of Tylosaurus has been suggested to being a weapon used by Tylosaurus against others of its kind during territorial combat. While only speculation, no ecosystem is capable of supporting a high number of large predators in the same area indefinitely, and it is at least possible that Tylosaurus may have been territorial and fought for the right to be active in an area where food was plentiful. For a long time,Tylosaurus was often reconstructed with a dorsal crest which was based upon fossils that appeared to show one running down the length of the animal. Later study revealed this to have been cartilage from the trachea, basically the windpipe that air passes down from the nostrils to the lungs. In life the cartilage would have kept the shape of the trachea preventing it from closing off when food was being swallowed just like in other animals. While it is rare for cartilage to be preserved, many sharks which have cartilaginous skeletons have been preserved on many occasions proving that when conditions are right it is possible for cartilage to be preserved, although with varying degrees of success. This is why accurate reconstructions omit the dorsal crest. Study of another mosasaur named Platecarpus has revealed that at least some mosasaurs may have had a tail fluke similar to the caudal (tail) fin of a shark. It is still a matter of debate if the same could be suggested for Tylosaurus, but some artistic renderings now include a vertical tail fluke.
Whereas Tyrannosaurus is dubbed “the land predator of the late Cretaceous”,Tylosaurus would have equally been “the predator of the late Cretaceous seas”, specifically the Western Interior Seaway that once submerged the central portion of the United States and Canada. The large size of Tylosaurus meant that it easily attained apex predator status and as such absolutely nothing was off the menu. Fossil evidence exists to support this as stomach contents for Tylosaurus are well-known and include fish, sharks, flightless birds like Hesperornis, large marine reptiles such as plesiosaurs and even other mosasaurs. There is also evidence to suggest that Tylosaurus fed upon dinosaurs, as evidenced by a specimen dubbed the “Talkeetna Mountains Hadrosaur” from the Matanuska Formation in Alaska. The bones associated with this hadrosaur have puncture marks that are very similar to the tooth size and arrangement, in fact detailed study has shown that a feeding Tylosaurus is the most likely cause of the marks. However this case of a Tylosaurus feeding upon a dinosaur is most likely a case of scavenging, with a Tylosaurus discovering the body of a dinosaur that had drowned and been swept out to sea. When animals die, gases given off by the decomposition processes build up inside the body causing it bloat and float up to the surface. Had a mosasaur fed from the carcass before this, the gashes and puncture wounds from the teeth would have caused the gases to escape causing the dinosaur to sink straight to the bottom rather than being washed out with the tides and currents. The carcass then seems to have sunk to the bottom through the release of the gas, either through natural decomposition, or more likely scavenging of other ocean predators such as sharks like Squalicorax or perhaps even the Tylosaurus itself. Study of the sea floor where the hadrosaur skeleton was found also shows that the largest concentration of flesh was on the side which was in contact with the ground. This shows that while the Tylosaurus was feeding from the carcass it was unable to reach the bottom side. If the Tylosaurus had attacked a living hadrosaur you would expect to see signs of flesh removal and tooth marks all over. The greatest number of tooth marks are on the outer extremities of the hadrosaur such as the lower legs which a mosasaur would have had an easier time manipulating rather than the bulk of the main body. Also the amount of flesh on the bone of the lower legs would have been much less, resulting in a higher incidence of bite marks as the teeth bit through the thin muscle and straight into the bone.
Although Tylosaurus had large teeth and powerful jaws, these may not have been the primary weapons of attack, at least not in the expected sense. The most forward areas of the jaws have a severe reduction in teeth to the point of being toothless. The snout is also reinforced making it stronger than other marine reptiles. Additionally some Tylosaurus snouts show signs of compression damage that seem to have been caused by a violent impact. These all point to Tylosaurus relying upon brute force to ram prey at high speeds. Such an attack probably would not kill prey outright, but could quite easily stun it so that it floated hopelessly in the water. It would have been particularly effective against other marine reptiles as they approached the surface to breathe air. Stunning them by ramming would not only make them a sitting target, but without a fresh supply of air they could have possibly drowned if Tylosaurus did not immediately return to finish its prey off. Despite the devastating nature of such an impact, such as attack would actually require patience and timing to execute properly, as while the prey was smaller, it was almost certainly more maneuverable than a fully grown Tylosaurus. This makes attacks in the same depth plane unlikely because despite its speed, prey would have likely seen the attack coming and taken avoiding action. This is why it makes the most sense for a marine predator to approach its target from below as not only can it home in unseen, other sensory adaptations such as the pressure and electro-receptors of fish tend to work in sideways directions, and any scent from the predator would also drift sideways in the ocean currents, not up. This style of attack has also been implied for the giant shark Megalodon as well as predatory whales such as Livyatan and suggests that while the predators and prey were different throughout the ages, the actual method of attack was principally the same. Smaller prey items like flightless birds such as Hesperornis and sea turtles probably did not require such special hunting strategies and may be cases of opportunistic feeding. The one drawback of being the biggest predator in the area is that you have to take every opportunity you can to feed or you risk starvation. Cases of Tylosaurus killing sharks and other mosasaurs could be presented as a case of intriguing predation. This is where a predator not only kills for food, but also removes potential competition from the ecosystem in the process.
#1 – MEGALODON
Phonetic: Meg-ah-low-don
Named By: Louis Agassiz - 1843
Synonyms: Procarcharodon megalodon, Megaselachus megalodon
Classification: Chordata, Chondrichthyes, Elasmobranchii, Lamniformes
Species: Carcharodon megalodon or Carcharocles megalodon, refer to text for full explanation
Diet: Carnivore
Size: Many estimates exist but usually between 15 and 17 meters long
Known locations: Worldwide
Time period: Late Oligocene to Early Pleistocene
Fossil representation: Mostly teeth, but some vertebrae are also known
Not only was Megalodon the biggest prehistoric shark that ever lived; it was the largest predatory marine creature in the history of the planet, outweighing both modern Great White Sharks and ancient reptiles like Liopleurodon and Kronosaurus. Megalodon didn’t earn its nickname (“Big Tooth”) for nothing. It was truly the “baddest of the bad” in prehistoric lore and without question, deserves the status as the #1-ranked Prehistoric Marine Predator. The teeth of this prehistoric shark were over half a foot long, serrated and heart-shaped (by comparison, the biggest teeth of a Great White Shark are only about 3 inches long). You have to go back 65 million years, to none other thanTyrannosaurus Rex to find a creature with consistently bigger choppers, though the canines of some Saber-toothed Cats also measured up. Because sharks are constantly shedding their teeth–thousands and thousands over the course of a lifetime–Megalodon teeth have been found all over the world, from antiquity to modern times. It was only in the 17th century that a court physician named Nicholas Steno identified peasant’s prized “tongue stones” as shark teeth; for this reason, some experts describe Steno as the world’s first paleontologist. In 2008, a joint research team from Australia and the U.S. used computer simulations to calculate Megalodon’s biting power. The results can only be described as terrifying: whereas a modern Great White Shark chomps with about 1.8 tons of force (and a lion with a wimpy 600 pounds), Megalodon chowed down on its prey with a force of between 10.8 and 18.2 tons–enough to crush the skull of a prehistoric whale as easily as a grape.












































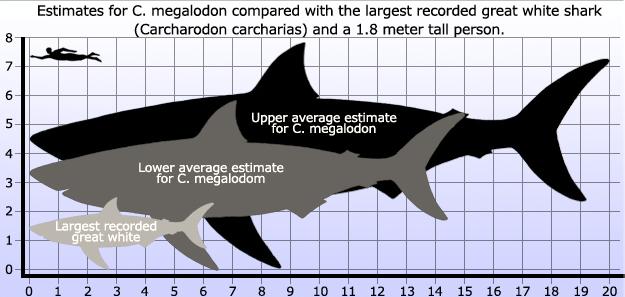
Since Megalodon is known from thousands of teeth but only a few scattered jaw fragments, its exact size has always been a matter of debate. Over the past century, paleontologists have come up with estimates (based mainly on tooth size and analogy with modern Great White Sharks) ranging from 40 to 100 feet, but the consensus today is that adults were 55 to 60 feet long and weighed as much as 100 tons and some superannuated individuals may have been even bigger. The ocean’s natural buoyancy allows “top predators” to grow to massive sizes, but none were more massive than Megalodon. The giant aquatic reptiles of the Mesozoic Era like Liopleurondon and Kronosaurus, “only” attained weights of 30 or 40 tons, and a modern Great White Shark can only aspire to a relatively puny 3 tons. The only marine animal to outclass Megalodon is the Blue Whale, individuals of which have been known to weigh well over 100 tons. Although the bigger than a Megalodon, the Blue Whale is technically a carnivore, it feeds mostly on tiny krill. Megalodon had a diet more befitting an apex predator, feasting on the prehistoric whales that swam the earth’s oceans during the Pliocene and Miocene epochs, but also chowing down on dolphins, squids, fish and even giant turtles whose shells as tough as they were, couldn’t hold up against 10 tons of biting force. Megalodon may even have attacked the giant whale Leviathan.
According to at least one analysis, Megalodon’s hunting style differed from that of modern Great White Sharks. Whereas Great Whites dive straight toward their prey’s soft tissue (say, a carelessly exposed underbelly), Megalodon’s teeth were suited to biting through tough cartilage and there’s some evidence that it may have first sheared off its victim’s fins (rendering it unable to swim away) before lunging in for the final kill. Technically, Megalodon is known as “Carcharodon megalodon”–meaning it’s a species (Megalodon) of a larger genus (Carcharodon). Also technically, the modern Great White Shark is known as “Carcharodon carcharias”, meaning it belongs to the same genus as Megalodon. However, not all paleontologists agree with this identification, claiming that Megalodon and the Great White arrived at their striking similarities via the process of convergent evolution. Unlike some marine predators of prehistoric times–which were restricted to the coastlines or inland rivers and lakes of certain continents–Megalodon had a truly global distribution, terrorizing whales in warm-water oceans all over the world. Apparently, the only thing keeping adult Megalodons from venturing too far toward solid land was their enormous size, which would have beached them as helplessly as a 16th-century Spanish Galleon. So Megalodon was huge, relentless and the apex predator of the Pliocene and Miocene epochs. What went wrong? Well, there’s no lack of theories: Megalodon may have been doomed by global cooling which culminated in the last Ice Age or by the gradual disappearance of the giant whales that constituted the bulk of its diet.